

Биомасса микроводорослей содержит множество биологически активных веществ, имеющих лекарственную и пищевую ценность, таких как полиненасыщенные жирные кислоты, каротиноиды, витамины и т. д., а биомасса микроводорослей также может использоваться для производства различных биоэнергий в качестве заменителя ископаемой энергии, например биоводорода. , биоэтанол и биодизель. Кроме того, микроводоросли являются не только естественной приманкой для животных аквакультуры, но и служат источником высококачественного белка для питания человека, решая проблемы продовольственной безопасности.

Однако крупномасштабное выращивание и коммерческое применение микроводорослей по-прежнему сталкивается с проблемами низкой фактической урожайности и высокой стоимости выращивания. Теоретическая максимальная эффективность использования солнечной энергии микроводорослями в процессе синтеза биомассы посредством фотосинтеза составляет 8%~10%, температура листьев составляет 30 °C, а концентрация CO2 составляет 387 мл/м3, C3 и C4. Теоретические максимумы скорости преобразования солнечной энергии для растений. составляют 4,6% и 6% соответственно. Однако в большинстве случаев фактическая скорость использования световой энергии при непрерывной культуре микроводорослей в лаборатории составляет всего около 3%, а степень конверсии крупномасштабной культуры еще ниже. Это указывает на то, что культура микроводорослей еще далека от реализации своего фотосинтетического потенциала и еще есть огромные возможности для оптимизации с точки зрения фотосинтетической способности связывать углерод и потенциала технологии выращивания.
1.Оптимизация стадий фотореакций в фотосинтезе микроводорослей
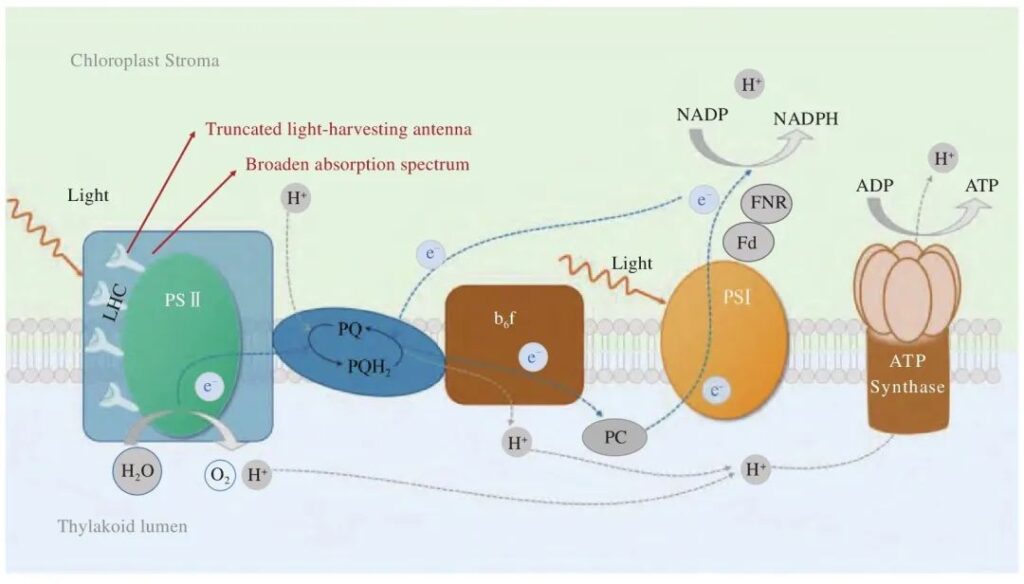
На рис. 1 представлена принципиальная схема стадий фотореакций микроводорослей. Подобно высшим растениям, микроводоросли захватывают световую энергию через различные типы светособирающих антенн (светособирающие комплексы) и проводят ее к реакционному центру фотосистемы II., где окисленная вода выделяет кислород, производит ионы водорода и электроны и в конечном итоге производит АТФ и НАДФН. Между фотосистемой II существует цепь переноса электронов. и I., который содержит акцепторы электронов, такие как пластидный хинон и цитохром b6f (Cyt b6f). Усечение светособирающей антенны и повышение эффективности использования световой энергии являются наиболее часто используемыми методами модификации стадии фотореакции микроводорослей.
2.Оптимизация темновой фазы реакции фотосинтеза микроводорослей
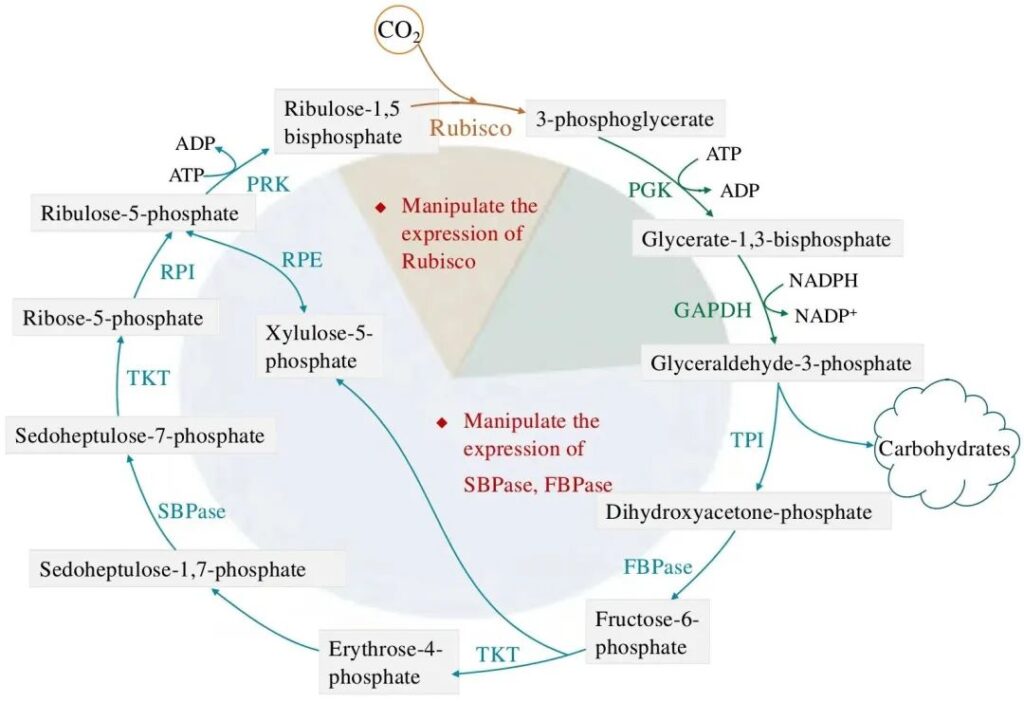
2.1 Модификация рибулозо-1,5-бисфосфаткарбоксилазы/оксигеназы
In the CBB cycle, ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) is responsible for catalyzing the reaction of 1 moleculeCO2 with 1 molecule Ribulose-1,5-bisphosphate (RuBP) to generate 2 molecules of 3-phosphoglycerate, which is considered to be the rate-limiting step of the entire cycle. Rubisco is the most abundant protein in the biosphere and forms a bridge between inorganic carbon and living organisms. But at the same time, Rubisco is also considered to be a very inefficient catalyst. Therefore, in higher plants, Rubisco has also been regarded as the primary target of photosynthetic modification, which has received great attention and research. The structure of Rubisco in cyanobacteria, diatoms, and green algae is very similar, consisting of 3 large subunits and 8 small subunits. The large and small subunit genes of the green algae Rubisco are present in the chloroplast and nuclear genomes, respectively, while the genes encoding the large and small subunits of diatoms are located in the chloroplast genomes. Microalgae have always been regarded as excellent “donors” of Rubisco, and the use of Rubisco as a source of microalgae to replace Rubisco in higher plants is regarded as a potential means to improve the photosynthesis efficiency and biomass of higher plants. However, due to the complex assembly mechanism of Rubisco, the heterologous expression of the microalgae Rubisco in plants did not achieve satisfactory results
2.2. Построение фотодыхательных ветвей.
Rubisco’s oxygenation produces the toxic metabolite 2-phosphoglycolic acid (2-PG), which needs to be recycled through the photorespiration pathway, but the photorespiration process requires the consumption of ATP and NADPH, and the release of immobilizedCO2 andNH3; 2 molecules of 2-PG are converted to 1 molecule of 3-phosphate glyceric acid (3-PGA) through the photorespiration pathway, and the whole process consists of distributed in chloroplasts, peroxisomes, Nine enzymatic steps in the mitochondria and cytoplasm are completed, and it is estimated that photorespiration can release up to 9/1 of the immobilizedCO4, resulting in a huge waste of C and N. However, the photorespiration process recovers 2% of C for cellular metabolism from 2 molecules 2-PG, which plays a very important role in cellular metabolism, therefore, theoretically, constructing a new photorespiration branch to avoid the waste ofCO75 andNH2 or increase the recovery capacity of releasingCO3 can improveCO2to increase biomass production rate. The feasibility of this scheme has been successfully demonstrated in the model organism Arabidopsis thaliana and the oil crop camelina, and the newly constructed photorespiratory branch can help the transformed strain significantly increase the biomass production rate.
2.3 Трансформация механизма обогащения углерода
В отличие от растений C3, большинство микроводорослей и растений C4 имеют механизм концентрации углерода (CCM), позволяющий справляться с более низкими концентрациями CO2 в водной среде. У цианобактерий Рубиско запечатан в карбоксисомах, тогда как у эукариотических микроводорослей Рубиско в основном присутствует в белковых ядрах хлоропластов, а концентрация CO2 вокруг активного центра Рубиско может более чем в 1000 раз превышать концентрацию CO2 в окружающей среде благодаря CCM. Карбоангидраза (КА) является ключевым ферментом, участвующим в CCM, и отвечает за катализацию обратимого взаимодействия между HCO3- и CO2. CCM в микроводорослях обычно представляет собой индуцируемый механизм, который регулирует уровень экспрессии CCM посредством восприятия окружающих концентраций CO2. При крупномасштабном выращивании микроводорослей часто используются концентрации CO2 значительно выше атмосферных, что может привести к отключению СКК.
Таким образом, многие важные новаторские успехи были достигнуты в трансформации и оптимизации фотосинтетических систем микроводорослей посредством усечения светособирающей антенны, увеличения светособирающей способности и использования световой энергии, преобразования цикла CBB и CCM, а также создания фотодыхательная ветвь. Однако общий прогресс отстает от трансформации и оптимизации путей фотосинтеза высших растений, особенно товарных культур, что приводит к нереализованному потенциалу связывания углерода микроводорослями. Благодаря добыче превосходных фотосинтетических элементов, регуляторных факторов и созданию новых путей можно эффективно снять ограничение на узкие места и реализовать энергетический баланс между световыми и темновыми реакциями. Фотосинтез не только регулируется светом, но и регулируется эффективностью потребления конечного продукта, что указывает на необходимость одновременного осуществления модификации метаболических путей, связанных с фотосинтезом, на клеточном уровне. Благодаря быстрому развитию синтетической биологии была заложена основа для разработки и создания высокоэффективных инженерных штаммов микроводорослей, улавливающих углерод, с использованием микроводорослей в качестве организмов-шасси для фотосинтетической секвестрации углерода. Методы и концепции синтетической биологии могут использоваться для разработки или синтеза фотосинтетических систем с более высокой эффективностью световой энергии и эффективностью связывания углерода, устойчивостью к фотоповреждениям и производством менее активных форм кислорода, независимо от видовых ограничений.
О БайЛун Биотехнологическая Компания, ООО:
БайЛун Biotechnology Co., Ltd. stands as a leading supplier and premier technical service provider, specializing in the provision of comprehensive bioreactor systems and advanced control solutions. Our extensive product line encompasses a wide array of offerings, ranging from bioreactors (fermenters) to animal cell bioreactors, biological shakers, and control systems tailored for bioprocessing applications. With a capacity spanning from 0.1L to 1000KL, we are committed to fostering the growth of China’s bioreactor industry on a global scale.
В БайЛун, we boast a seasoned team of engineers equipped with profound expertise in fermentation processes, biochemical equipment, and chemical technology. Moreover, we actively engage nationally renowned experts and scholars as technical consultants, ensuring the robust technological underpinnings of our products. Central to our ethos is a relentless pursuit of product innovation and technological leadership, all geared towards guaranteeing utmost customer satisfaction. We hold ourselves accountable to prioritize customer benefits, embodying this commitment as the cornerstone of Bailun Company’s core values.
Наш разнообразный портфель продуктов тщательно разработан для удовлетворения разнообразных потребностей наших клиентов. С самого начала мы придерживались принципов приоритета качества, клиентоориентированности и честности операций. Наша непоколебимая приверженность удовлетворению растущих потребностей наших клиентов двигает нас вперед. Принимая волну экономической глобализации, мы искренне приглашаем к сотрудничеству предприятия по всему миру, стремясь к взаимному процветанию и успеху.
Дух БайЛун воплощает в себе приверженность клиентоориентированности, стремление к совершенству в области качества, приверженность справедливости и порядочности, а также непоколебимое стремление к постоянному совершенствованию и инновациям.
Связаться с нами:
Добавить: 6848# Liuxiang Rd., Цзядин, Шанхай, Китай
Контактное лицо: Мэдди
Электронная почта: sales@fermentertech.com
Телефон:+86-134-7276-8163 (WhatsApp)
Веб-сайт: http://fermenterchina.com/